- Return to Article Of The Month index
Nutritional Strategies for Developing Bos Indicus-Influenced Beef Heifers
February, 2020
Philipe Moriel, UF/IFAS Range Cattle Research & Education Center, Ona
Introduction
A
major limiting factor for reproductive success of Bos indicus-influenced
beef heifers is the late attainment of puberty due to genetics, environment
(i.e. heat stress), and nutrition. Heat stress is detrimental to cattle
metabolism, growth, reproduction, health, and welfare (Mader, 2003; Key et
al., 2014) and will become a greater challenge in the future due to the
potential impact of global climate change (IPCC, 2007). Environmental
conditions are considered thermoneutral when thermal-humidity index (THI)
≤ 70, mild heat stress when 70 ≤ THI < 74, heat stress when 74 ≤ THI < 77,
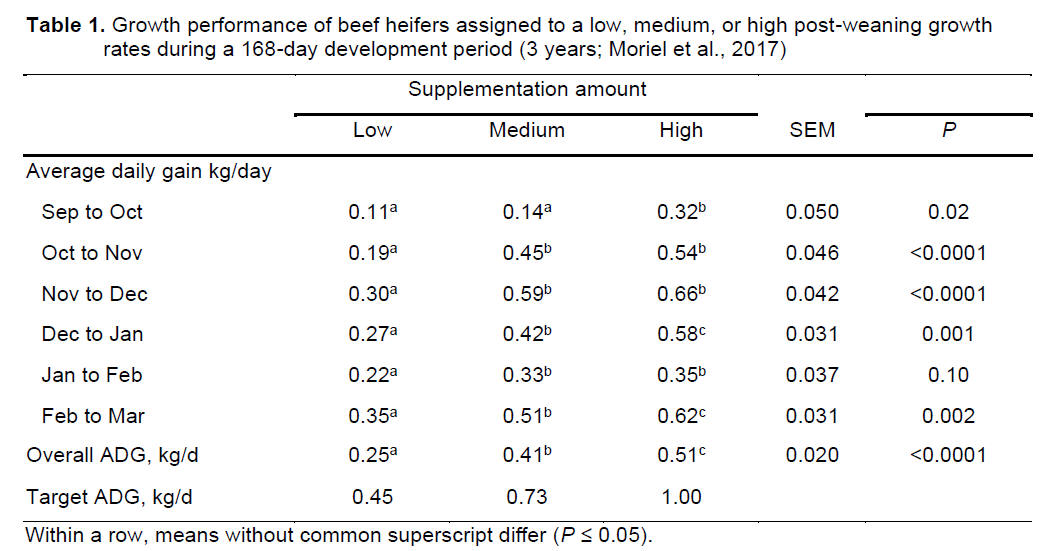
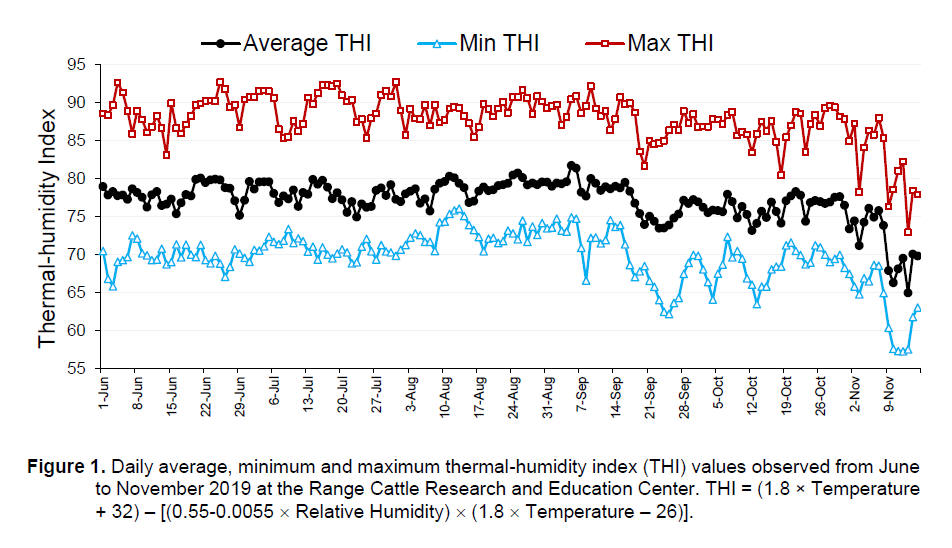
and severe heat stress when THI ≥ 77 (Davis et al., 2003). Figure 1 shows
the average, minimum and maximum daily THI values obtained at the University
of Florida - Range Cattle Research & Education Center (Ona, FL). From June
to October 2019, average THI values were within or above the threshold
considered as heat stress. Also, maximum THI values often reached severe
heat stress levels. These challenging conditions during summer partially
explain the poorer average daily gain (ADG;
Table 1)
of heifers, despite the greater nutritional composition of forage during
Summer vs. Fall.
The cow-calf industry in Florida relies on
warm-season forages as the main source of feed for beef cattle. This forage
type often do not meet the requirements of growing heifers, even if herbage
mass is not a limiting factor. Moore et al. (1991) compiled the nutritional
analysis of 637 samples of forages commonly grown in Florida (bahiagrass,
bermudagrass, digitgrass, stargrass, and limpograss) and reported that most
of these grasses contained between 5 to 7% crude protein (CP) and 48
to 51% total digestible nutrients (TDN), on the basis of dry matter (DM).
Developing heifers require diets with at least 55% TDN and 8.5% CP on a DM
basis to achieve adequate growth rates (≥ 0.50 kg/d; NRC, 1996).
Nevertheless, successful reproductive performance can still be obtained if
heifers become pubertal before the initiation of breeding season (Moriel et
al., 2017). For instance, our previous study funded by the Florida Cattlemen
Enhancement Board demonstrated that average final pregnancy rates were 82%
for heifers that achieved puberty BEFORE the start of the breeding season
compared to 36% for heifers that achieved puberty DURING the breeding
season. In this article, we will provide a summary of our previous and
on-going studies to optimize growth and reproduction of Bos indicus-influenced
beef heifers in tropical/subtropical environments.
Post-Weaning Energy Intake
Frequency of Concentrate Supplementation
Previous studies reported that reducing the
frequency of energy supplementation from daily to 3 times weekly had no
impact (Drewnoski et al., 2011; Moriel et al., 2016) or decreased ADG of
beef calves by 10 to 21% (Cooke et al., 2008; Artioli et al., 2015).
Discrepancies among these results can be associated to differences in
supplement composition, animal breed and sex, location of the study, forage
species and quality, and the potential interactions among those factors
(Artioli et al., 2015). However, differences in daily forage DM intake
between cattle offered frequent or infrequent energy supplementation is the
primary factor explaining the variable growth performance among these
studies. When supplementation frequency is reduced, cattle
consume a large portion of concentrate in a single day and receive no
concentrate supplementation on the next day. This less frequent schedule of
supplementation leads to fluctuations in forage and nutrient intake.
In terms of performance of beef heifers, reducing
the frequency of supplementation may be detrimental to reproduction.
Moriel et al. (2012) evaluated the impact of similar weekly energy
supplementation that was offered either daily (S7 heifers) or 3 times
weekly (S3 heifers; Monday, Wednesday, and Friday) on growth and
reproductive performance of developing beef heifers fed stargrass.
Supplements were offered at weekly rates of 16 kg of DM/heifer. On days that
both S3 and S7 heifers were supplemented, S3 heifers had lower hay DM intake
compared with S7 heifers (2.55 vs. 3.36 kg/day, respectively). On days that
only S7 heifers were supplemented, S3 heifers also had lower hay DM intake
(3.15 vs. 3.38 kg/day for S3 and S7 heifers, respectively). Consequently,
overall mean hay DM intake was 15.4% lower for S3 vs. S7 heifers (2.85 vs.
3.37 kg/day, respectively). Estimated NEg intake followed the same pattern
observed on total DM intake, and overall estimated NEg intake was slightly
greater for S7 vs. S3 heifers (2.75 vs. 2.59 Mcal/day, respectively).
However, the magnitude of differences on estimated overall NEg intake
between S7 and S3 heifers was not sufficient to impact ADG (0.28 vs. 0.27
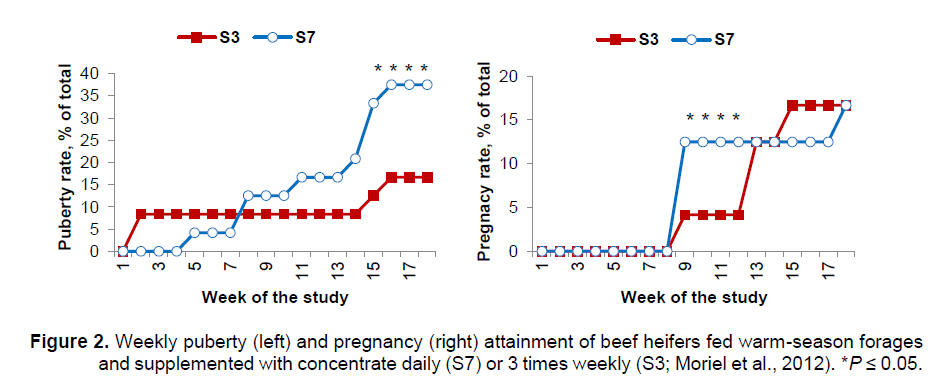
kg/day for S3 and S7, respectively). Despite the similar ADG, attainment of
puberty and pregnancy were delayed by decreasing the frequency of energy
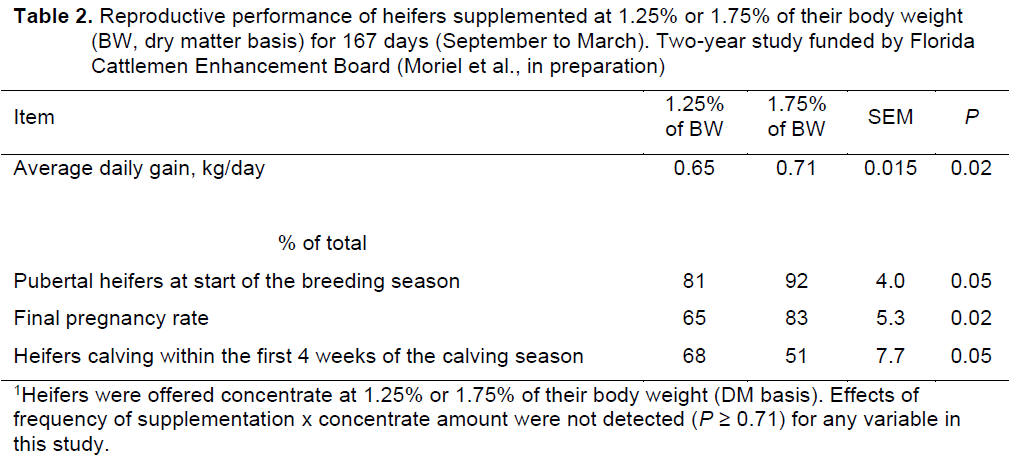
supplementation (Figure 2). At the end of the breeding season,
approximately 38% of S7 heifers were pubertal, whereas only 17% of S3
heifers were pubertal. Final pregnancy rates did not differ between
treatments but S7 heifers became pregnant earlier in the breeding season (Figure
2).
Enhanced reproductive performance have been
associated with increased blood concentrations of glucose, insulin, and
insulin-like growth factor 1 (IGF-1; Hess et al., 2005). In cattle,
GnRH secretion is impaired when glucose availability is inadequate, but
resumed when glucose levels are adequate (Hess et al., 2005). Cows with low
plasma insulin concentrations have impaired LH surge, reduced numbers of LH
receptors in the dominant follicle, and fail to ovulate (Diskin et al.,
2003). Insulin-like growth factor 1 is a major metabolic signal regulating
reproduction in cattle (Wettemann and Bossis, 2000; Thatcher et al., 2001).
Plasma glucose, insulin and IGF-1 are positively affected by nutrient intake
(Vizcarra et al., 1998; Bossis et al., 1999) and supplementation frequency
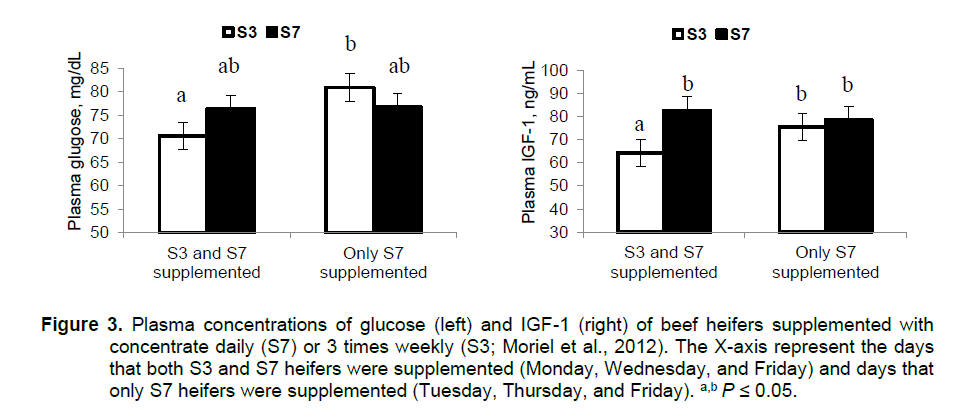
(Cooke et al., 2007). For instance, plasma glucose and insulin
concentrations were greater for S3 vs. S7 heifers on the days that only S7
heifers received supplementation, but not on days that both treatment groups
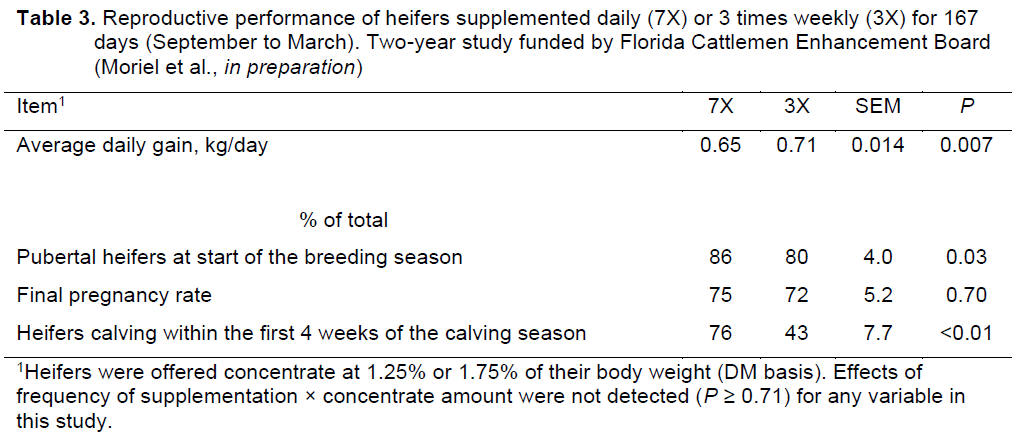
were supplemented. More importantly, heifers supplemented every day had less
daily variation in plasma concentrations of glucose and IGF-I than heifers
supplemented 3 times weekly (Figure 3; Moriel et al., 2012). The differences
in plasma concentrations of glucose and insulin were attributed to the
pattern of nutrient intake of each treatment, and this lower fluctuation in
blood parameters with a more frequent supplementation schedule likely
collaborated for the improved puberty achievement compared to infrequent
supplementation (Moriel et al., 2012).
Recently, we attempted to overcome the negative
effects of frequency of supplementation by increasing the amount of
supplement offered to heifers. In this 2-year study, heifers were
supplemented with concentrate DM at: 1.25% of body weight offered 3 times
weekly (1.25-3X); 1.25% of body weight offered 7 times weekly (1.25-7X);
1.75% of body weight offered 3 times weekly (1.75-3X); or 1.75% of
body weight offered 7 times weekly (1.75-7X). The hypothesis was that
by increasing the concentrate supplementation amount, heifers offered
reduced frequency of supplementation would achieve similar puberty
attainment and pregnancy percentage compared to heifers supplemented daily.
Contrary to our hypothesis, effects of supplementation frequency × amount
were not detected (P ≥ 0.71) for any variable. Growth and
reproductive performance of heifers supplemented at 1.25% or 1.75% were
discussed previously (Table 2). Similar to our previous studies, growth and
reproductive performance of heifers supplemented 3 times weekly were reduced
compared to heifers supplemented daily (Table 3). Although pregnancy rates
did not differ, heifers supplemented 3 times weekly calved later during
their first calving season compared to heifers supplemented daily (Table 3).
Therefore, despite including greater supplementation amounts and a puberty
induction protocol, heifer reproductive performance was significantly
jeopardized when supplementation frequency was reduced from daily to 3 times
weekly.
Growth Pattern (Stair-Step Strategy)
The experiment will be conducted at the Range Cattle
REC (Ona, FL) from September 2019 to June 2020 (Year 1) and replicated from
September 2020 to June 2021 (Year 2). In September of each year, 64 Brangus
heifers will be allocated into 1 of 16 bahiagrass pastures (4
heifers/pasture). Treatments will be assigned to pastures (8
pastures/treatment) and will consist of: control heifers supplemented with
concentrate DM at 1.50% of body weight from September until the start of the
breeding season in December (day 0 to 100 of the study; CON); or
stair-step heifers initially offered concentrate DM at 1.05% of body weight
from September to October (day 0 to 50 of the study), and then,
concentrate DM at 1.95% of body weight (DM basis) from October until the
start of the breeding season in December (SST; day 50 to 100 of the
study). In average, both treatments will be supplemented with concentrate DM
at 1.50% of body weight from September to December (22% CP and 73% TDN; DM
basis).
In year 1, total supplement DM offered to heifers
did not differ between treatments (410 vs. 405 ± 3.5 kg/heifer for SST and
CON, respectively; P = 0.26). In terms of growth, ADG from day 0 to
50 did not differ between treatments (0.63 vs. 0.62 ± 0.040 kg/day; P
= 0.87) but was greater for SST vs. CON heifers from day 50 to 100 (0.73 vs.
0.56 ± 0.044 kg/day; P = 0.01), leading to a tendency for greater
overall ADG (0.68 vs. 0.59 ± 0.031 kg/day; P = 0.07) and greater body
weight at start of estrus synchronization protocol for SST vs. CON heifers
(311 vs. 302 ± 2.1 kg; P = 0.009).
Intravaginal thermometers were inserted into heifers
to determine the intravaginal temperatures every 30 min from day 25 to 31
(Sep 7th to 12th) and day 85 to 91 of the study (Nov 6th
to 12th; see Figure 1 for THI values). From day 25 to 31, SST
heifers had significantly lower intravaginal temperatures from 0930 h to
1800 h compared to CON heifers (nearly 0.25 to 0.32ºC lower for SST vs.
CON), which is likely a result of lower heat increment and partially
explains the lack of treatment effects on heifer ADG from day 0 to 50
despite the drastic differences in supplement DM offered (1.05 vs. 1.50% of
body weight for SST and CON, respectively). From day 85 to 91, supplement DM
amount did not (P = 0.39) affect intravaginal temperature of heifers,
which likely prevented energy waste to cope with heat stress and allowed the
greater ADG of SST vs. CON heifers.
Although overall ADG tended to differ, reproductive
tract scores (4.52 vs. 4.37 ± 0.173 for SST and CON, respectively; P
= 0.58) and percentage of pubertal heifers at the start of the
synchronization protocol (79.3 vs. 71.9 ± 8.23 % of total for SST and CON,
respectively; P = 0.54) did not differ between treatments. We will
repeat this study for another year to confirm these results, but based on
data from year 1, the SST strategy offered an opportunity to harvest greater
growth performance before the start of the breeding season without
increasing feed costs. This enhanced growth performance did not lead to any
advantage on heifer puberty attainment before breeding in year 1 of our
study but might be important in situations when heifer post-weaning body
weight are lighter than those reported herein.
Early-Weaning
In January of each year (day 0 of the study),
Brangus calves (70 days of age) were assigned to remain with their dams and
be normally weaned at 250 days of age (day 180 of the study; NW), or
early-weaned at 70 days of age and randomly assigned to 1 of 3 early-weaning
management systems from day 0 to 180 of the study: 1) ryegrass and
bahiagrass grazing for 180 days (EWPAST); 2) high-concentrate diet in
drylot for 180 days (EW180); and 3) high-concentrate diet in drylot
for 90 days, then bahiagrass grazing for additional 90 days (EW90).
When early-weaned calves were in drylot, they were limit-fed the
high-concentrate diet at 3.5% of body weight (as-fed). When early-weaned
calves were on pasture, they were supplemented with the same
high-concentrate diet at 1.0% of body weight (as-fed). Calves that were kept
with the mothers until weaning (250 days of age) did not receive
supplementation from 70 to 250 days of age.
We observed that EW90, EW180, and EWPAST heifers had
similar or greater growth performance from day 0 to 180 than NW heifers
(Table 4). From day 180 of the study until the end of the breeding season
(day 395), all heifers were supplemented with concentrate DM at 1.5% of body
weight (as-fed). During this period, no differences were detected for ADG
among treatments (in average = 0.68 kg/day). Interestingly, limit-feeding a
high-concentrate diet in drylot, for at least 90 days, increased the
percentage of heifers cycling at the start of the breeding season compared
to normally weaned heifers (Table 4). More specifically, a greater
percentage of early-weaned heifers fed high-concentrate diet in drylot for
only 90 days achieved puberty at the start of the breeding season, despite
having similar body weight and ADG compared NW heifers. This response
indicates that we can successfully hasten puberty achievement if Bos
indicus-influenced beef heifers by temporarily exposing young calves to
high-concentrate diets and high-growth rates starting at approximately 70
days of age.
Pre-Weaning Injections of Bovine Somatotropin
Bos taurus
and
Bos indicus
are different subspecies that
diverge in social and biological functions (Cooke et al., 2020).
Under the same environmental and nutritional
conditions, Bos taurus and Bos indicus cattle not only exhibit
diet-dependent differences in intake, digestion and ruminal fermentation (Habib
et al., 2008; Bell et al., 2017), but also
different ovarian function, circulating hormones and metabolites
(Sartori et al., 2016). These differences may determine the direction and
magnitude of performance responses to similar management applied to
Bos taurus
or Bos indicus breeds. Thus, we conducted 2 studies to
evaluate the impacts of preweaning injections of bST on growth and
reproductive performance of Brangus (Bos indicus × taurus;
Experiment 1; Piccolo et al., 2018) and Nellore beef heifers (Bos indicus;
Experiment 2; Moriel et al., 2019).
In Experiment 1, suckling Brangus heifers were
stratified by body weight (147 ± 20 kg) and age (134 ± 11 days) on day 0,
and randomly assigned to receive an s.c. injection of saline (SAL; 5
mL; 0.9% NaCl) or 250 mg of sometribove zinc (BST; Posilac, Elanco,
Greenfield, IN) on days 0, 14, and 28. Heifers and respective dams were
managed as a single group on bahiagrass pastures from day 0 until weaning
(day 127), and provided the same diet during the entire post-weaning phase.
In Experiment 2, suckling Nellore heifers were stratified by body weight (97
± 16 kg) and age (80 ± 10 days), and randomly assigned to receive s.c.
injections of saline (5 mL 0.9% NaCl) or 250 mg of sometribove zinc (BST)
on days 0 and 10 of the study. Then, all Nellore heifers were managed as a
single group in Brachiaria decumbens pastures, weaned on day 177, and
provided a corn silage–based TMR from weaning until the end of the study
(day 380).
In Experiment 1, Brangus-crossbred heifers
administered preweaning bST injections had an 8.6 ng/mL increase in plasma
IGF-1 concentrations (103 vs. 95 ± 3.2 ng/mL; P = 0.05) and 7.2%
increase on ADG from days 0 to 42 (1.15 vs. 1.07 ± 0.03 kg; P =
0.07), but no differences on overall pre-weaning ADG (0.88 and 0.89 ± 0.02
kg/day; P = 0.50) and post-weaning ADG (0.28 and 0.30 ± 0.02 kg/day;
P = 0.61) compared to saline heifers. Also, heifers assigned to BST
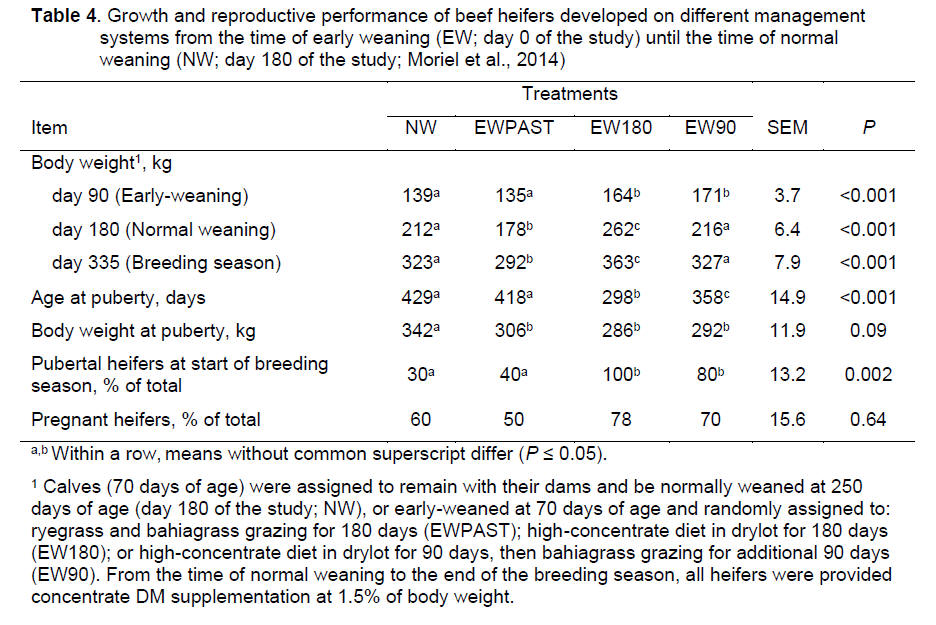
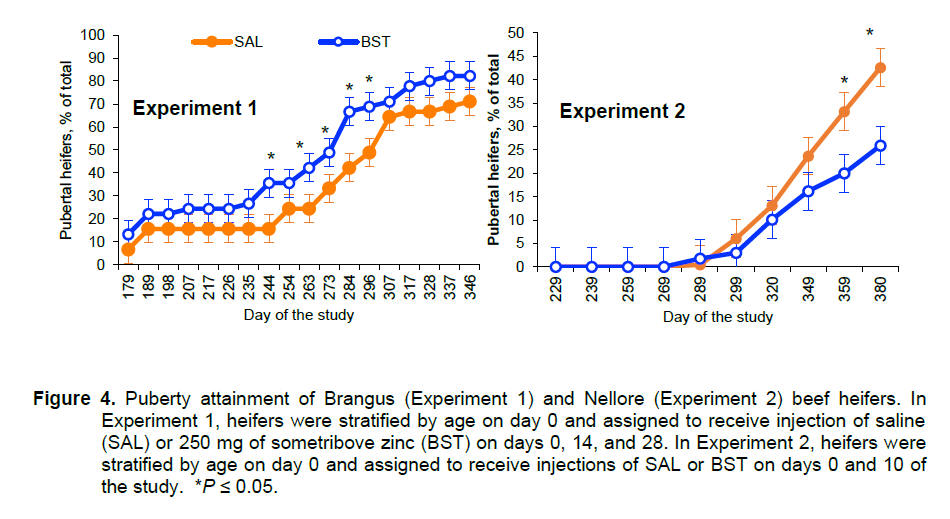
tended to achieve puberty 26 days earlier (388 vs. 414 ± 13 days; P =
0.10), had greater percentage of pubertal heifers on days 244, 263, 284, and
296 of the study (P ≤ 0.04; Fig. 4), and
tended to have greater overall pregnancy percentage (82 vs. 69 ± 6.1%; P
= 0.10) compared to saline heifers.
In Experiment 2, preweaning bST injections increased
plasma IGF-1 concentrations by 52 ng/mL (211 vs. 159 ± 9.3 ng/mL; P =
0.0001) and ADG from days 0 to 10 by 35% (0.65 vs. 0.48 ± 0.061 kg/day; P
= 0.03), but did not affect overall pre-weaning ADG (0.45 vs. 0.47 ± 0.009
kg/day; P = 0.24), tended to decrease post-weaning ADG by 3.6% (0.80
vs. 0.83 ± 0.014 kg/day; P = 0.07) and decreased puberty attainment
on days 349, 359, and 380 (P ≤ 0.05; Fig. 4) compared to saline
injections.
Sartori et al. (2016) reported that B. indicus cattle naturally have
greater circulating IGF-I concentrations compared with B. taurus
cohorts. Moreover, Mendonça et al. (2013) demonstrated that even under the
same environment and diet, Bos taurus–influenced dairy cows have less
circulating concentrations of IGF-I compared to Bos indicus cows,
which might be related to the different organ sensitivity to IGF-1. It is
possible that the greater increment on plasma IGF-1 concentrations following
bST injection in Experiment 2 vs. 1, in combination with the interval
between bST injections, was detrimental to the development of the
reproductive axis of Nellore heifers. Further studies investigating the
effects of breed on ovarian activity and gene expression in reproductive
tissue organs and brain, following bST injections, are warranted to confirm
this hypothesis.
Conclusions
Despite the challenges encountered by Bos indicus-influenced
beef heifers including extreme heat and humid conditions in combination with
forages of relatively poor nutritional composition, acceptable reproductive
performance may still be achieved. Some of these successful nutritional
management practices to enhance growth and reproduction included: increasing
the concentrate DM offered to heifers from 1.25% to 1.75% of body weight;
daily rather than infrequent (3X/week) concentrate supplementation;
stair-step strategy to boost growth (reproductive performance to be tested
in 2019/2020); and early-exposure to high-concentrate diets. Although
preweaning injections of bST are currently not allowed for beef cattle, our
results indicated that early manipulation of the somatotrophic axis may
benefit the reproductive performance of Brangus but not Nellore beef
heifers. Identifying additional strategies that can enhance calf performance
during early postnatal life may provide unique opportunities to optimize
feed resources and increase the profitability of beef cattle operations.
References
Artioli, L. F. A., P. Moriel, M. H. Poore, R. S.
Marques, and R. F. Cooke. 2015. Decreasing the frequency of energy
supplementation from daily to three times weekly impairs growth and humoral
immune response of preconditioning beef steers. J. Anim. Sci. 93:5430-5441.
Bell, N. L., R. C. Anderson, T. R. Callaway, M. O.
Franco, J. E. Sawyer, T. A. Wickersham. 2017. Effect of monensin inclusion
on intake, digestion, and ruminal fermentation parameters by Bos taurus
indicus and Bos taurus taurus steers consuming bermudagrass hay.
J. Anim. Sci. 95:2736–2746.
Bossis, I., R. P. Wettemann, S. D. Welty, J. A.
Vizcarra, L. J. Spicer, and M. G. Diskin. 1999. Nutritionally induced
anovulation in beef heifers: ovarian and endocrine function preceding
cessation of ovulation. J. Anim. Sci. 77:1536-1546.
Cooke, R. F., J. D. Arthington, C. R. Staples, W. W.
Thatcher, and G. C. Lamb. 2007. Effects of supplement type on performance,
reproductive, and physiological responses of Brahman-crossbred females. J.
Anim. Sci. 85:2564-2574.
Cooke, R. F., J. D. Arthington, D. B. Araujo, G. C.
Lamb, and A. D. Ealy. 2008. Effects of supplementation frequency on
performance, reproductive, and metabolic responses of Brahman-crossbred
females. J. Anim. Sci. 86:2296–2309.
Cooke, R. F., D. W. Bohnert, C. L. Francisco, R. S.
Marques, C. J. Mueller, and D. H. Keisler. 2013. Effects of bovine
somatotropin administration on growth, physiological, and reproductive
responses of replacement beef heifers. J. Anim. Sci. 91:2894–2901.
Cooke, R. F., C. L. Daigle, P. Moriel, S. B. Smith,
L. O. Tedeschi, and J. M. B. Vendramini. 2020. Board Invited Review - Cattle
adapted to tropical and subtropical environments (I): social, nutritional,
and carcass quality considerations. J. Anim. Sci. in press
doi: 10.1093/jas/skaa014.
Cushman, R. A., L. K. Kill, R. N. Funston, E. M.
Mousel, and G. A. Perry. 2013. Heifer calving date positively influences
calf weaning weights through six parturitions. J. Anim. Sci.
2013.91:4486–449.
Cushman, R. A., A. K. McNeel, and H. C. Freetly.
2014. The impact of cow nutrient status during the second and third
trimesters on age at puberty, antral follicle count, and fertility of
daughters. Livest. Sci. 162:252–258.
Davis, M. S., T. L. Mader, S. M. Holt, and A. M.
Parkhurst. 2003. Strategies to reduce feedlot cattle heat stress: Effects on
tympanic temperature. J. Anim. Sci. 2003. 81:649–661.
Diskin, M. G., D. R. Mackey, J. F. Roche, and J. M.
Sreenan. 2003. Effects of nutrition and metabolic status on circulating
hormones and ovarian follicle development in cattle. Anim. Reprod. Sci.
78:345-370.
Freetly, H. C., K. A. Vonnahme, A. K. McNeel, L. E. Camacho, O. L. Amundson,
E. D. Forbes, C. A. Lents, and R. A. Cushman. 2014. The consequence of level
of nutrition on heifer ovarian and mammary development. J. Anim. Sci.
92:5437–5443
Habib M. Pollott G. E. Leaver J. D. 2008. Effect of
cattle genotype and variable feed supply on forage intake and digestibility.
Asian-Australas. J. Anim. Sci. 21:1435–1440.
Hess, B. W., S. L. Lake, E. J. Scholljegerdes, T. R.
Weston, V. Nayigihugu, J. D. C. Molle, and G. E. Moss. 2005. Nutritional
controls of beef cow reproduction. J. Anim Sci. 83:E90-106E.
Horn, G. W., and F. T. McCollum. 1987. Energy
supplementation of grazing ruminants. Pages 125–136 in Proc. Graz. Livest.
Nutr. Conf., Jackson Hole, WY.
IPCC (Intergovernmental Panel on Climate Change:
AR4). 2007. The Intergovernmental Panel on Climate Change 4th Assessment
Report. Accessed May 12, 2011. http://www.ipcc.
ch/ Jackson Institute.
Key, N., S. Sneeringer, and D. Marquardt. 2014.
Climate change, heat stress, and U.S. dairy production. USDA Economic
Research Report #175. Social Science Research Network.
doi:10.2139/ssrn.2506668
Lucas,
A. 1991. Programming by early nutrition in man. Ciba Found. Symp.156:38-50.
Lynch, J. M., G. C. Lamb, B. L. Miller, R. T.
Brandt, Jr, R. C. Cochran, and J. E. Minton. 1997. Influence of timing of
gain on growth and reproductive performance of beef replacement heifers. J.
Anim. Sci. 75:1715–1722.
Mader, T. L. 2003. Environmental stress in confined
beef cattle. J. Anim. Sci. 81(E. Suppl. 2):E110–E119
Martin, C., L. Millet, G. Fonty, and B.
Michalet-Doreau. 2001. Cereal supplementation modified the fibrolytic
activity but not the structure of the cellulolytic bacterial community
associated with rumen solid digesta. Reprod. Nutr. Dev. 41:413–424.
Mendonça, L. G., N. B. Litherland, M. C. Lucy, D.
H. Keisler, M. A. Ballou, L. B. Hansen, and R. C. Chebel. 2013. Comparison
of innate immune responses and somatotropic axis components of Holstein and
montbéliarde-sired crossbred dairy cows during the transition period. J.
Dairy Sci. 96:3588–3598. doi:10.3168/jds.2012-5804
Miller, A. J., D. B. Faulkner, R. K. Knipe, D. R.
Strohbehn, D. F. Parrett, and L. L. Berger. 2001. Critical control points
for profitability in the cow-calf enterprise. Prof. Anim. Sci. 17:295-302.
Moore, J. E., W. E. Kunkle, and W. F. Brown. 1991. Forage quality and the
need for protein and energy supplements. pp. 113-123 in 40th Annual Florida
Beef Cattle Short Course Proceedings, Univ. of Florida, Gainesville.
Moriel, P., R. F. Cooke, D. W. Bohnert, J. M. B.
Vendramini, and J. D. Arthington. 2012. Effects of energy supplementation
frequency and forage quality on performance, reproductive, and physiological
responses of replacement beef heifers. J. Anim. Sci. 90:2371-2380.
Moriel,
P., S. E. Johnson, J. M. B. Vendramini, V. R. G. Mercadante, M. J. Hersom,
and J. D. Arthington. 2014 Effects of metabolic imprinting and calf
management systems on growth and reproductive performance of beef heifers.
J. Anim. Sci. 92:3096-3107.
Moriel, P., L. F. A. Artioli, M. B. Piccolo, M. H.
Poore, R. S. Marques, and R. F. Cooke. 2016. Decreasing the frequency and
rate of wet brewer’s grains supplementation did not impact growth but
reduced humoral immune response of preconditioning beef heifers J. Anim.
Sci. 94:3030-3041.
Moriel, P., Lancaster, P., G. C. Lamb, J. M. B. Vendramini, and J. D.
Arthington. 2017. Effects of post-weaning growth rate and puberty induction
protocol on reproductive performance of Bos indicus-influenced beef heifers.
J. Anim. Sci. 95:3523-3531.
Moriel, P., B. I. Cappellozza, M. B. Piccolo, R. F. Cooke, M. F. Miranda, L.
F. D. Batista, R. S. Carvalho, E. A. Colombo, F. V. Santili, R. V. O. Filho,
V. S. M. Ferreira, and J. L. M. Vasconcelos.
2019. Pre- and post-weaning injections of bovine somatotropin to optimize
puberty achievement of Bos indicus beef heifers. Trans. Anim. Sci. 3:
443-455.
NRC. 1996. Nutrient Requirements of Beef Cattle. 7th
rev. ed. National Academy Press, Washington, DC.
Piccolo, M. B., J. D. Arthington, G. M. Silva, G. C.
Lamb, R. F. Cooke, and P. Moriel. 2018. Pre-weaning injections of bovine ST
enhanced reproductive performance of Bos indicus-influenced replacement beef
heifers. J. Anim. Sci. 96:618-631.
Sartori, R., L. U. Gimenes, P. L. J. Monteiro Jr, L. F. Melo, P. S.
Baruselli, and M. R. Bastos.
2016. Metabolic and endocrine differences between Bos taurus and Bos indicus
females that impact the interaction of nutrition with reproduction.
Theriogenology 86:32–40
Schillo, K. K., J. B. Hall, and S. M. Hileman. 1992. Effects of nutrition
and season on the onset of puberty in the beef heifer. J. Anim. Sci.
70:3994–4005.
Spicer, L. J., and S. E. Echternkamp. 1995. The
ovarian insulin and insulin-like growth factor system with an emphasis on
domestic animals. Domest. Anim. Endocrinol. 12:223–245.
Thatcher, W. W., A. Guzeloglu, R. Mattos, M.
Binelli, T. R. Hansen, and J. K. Pru. 2001. Uterine-conceptus interactions
and reproductive failure in cattle. Theriogenology 56:1435–1450.
Vizcarra, J. A., R. P. Wettemann, J. C. Spitzer, and
D. G. Morrison. 1998. Body condition at parturition and postpartum weight
gain influence luteal activity and concentrations of glucose, insulin, and
nonesterified fatty acids in plasma of primiparous beef cows. J. Anim. Sci.
76:927-936.